Together, the processes of photosynthesis and cellular respiration allow life on Earth to gather energy for use in other reactions. Besides the organisms that rely on sulfur near hydrothermal vents, the majority of life on Earth relies on the sugar glucose. Glucose is created by the process of photosynthesis. Cellular respiration involves the breakdown of glucose and the storage of the energy received into the molecule ATP. Plants create their own energy through photosynthesis and also use cellular respiration to produce ATP. Animals must rely on the sugars that they’ve gathered from plants to supply their mitochondria material to produce ATP.
Process of Photosynthesis
Photosynthesis is the main process which drives life on Earth. Through photosynthesis, energy from the sun is captured in the bonds of organic molecules. These molecules, glucose molecules, are the basis of all life on Earth. Glucose will be used by the process of cellular respiration to harness chemical energy stored within the covalent bonds of the sugar.
Photosynthesis occurs in the leaves and green parts of plants. Organelles within plant cells, known as chloroplasts, contain specialized proteins capable of interacting with light. Cytochromes are these specialized proteins, which are attached to a heme group. Heme groups are also seen bound to hemoglobin, in blood cells. Instead of iron, these heme cells bind magnesium. The complex structure of the heme interacts with the photons of light passing through them.
The chloroplast uses the energy harnessed from these photons and their interaction with the cytochromes and other proteins to drive the formation of glucose. To do this, the chloroplasts will combine units of carbon dioxide into chains of 6 carbons, 12 hydrogens, and 6 oxygens. This is glucose, which can then be modified and combined with other glucose molecules to be stored as starches and complex sugars like fructose.
Photosynthesis Reaction
The photosynthesis reaction has two parts, commonly referred to as the Light reactions and the Calvin Cycle. The entire process of photosynthesis can be seen below.
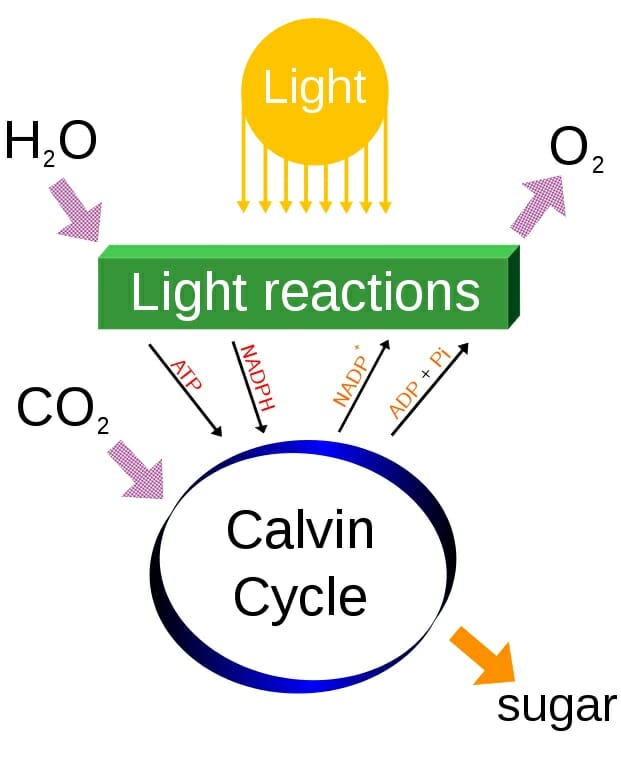
Simple photosynthesis overview
At the top of the diagram, light and water combine in the chloroplasts, where the hydrogens are separated from the oxygen in chain of proteins starting from the energy-collecting cytochromes and accessory pigments. The hydrogens, electrons, and associated energy are bound to ADP and NADP+. These molecules can bind a hydrogen, electrons, and energy. In doing so, they become the main products of the light reactions, NADPH and ATP. Oxygen is produced as a by-product.
ATP and NADPH are then used within the Calvin Cycle, a series of reactions which recycles these electron-carriers and produces glucose. The energy within and the hydrogen molecules are used to energize reactions throughout the cycle. The Calvin Cycle has three phases, carbon fixation, reduction, and regeneration of ribose. These reactions can be seen in the image below. Notice that the addition of one carbon dioxide in one turn of the reaction produces the 3-carbon molecule 3-phoshphoglycerate. Two of these molecules are then combined to produce a glucose, among other things.
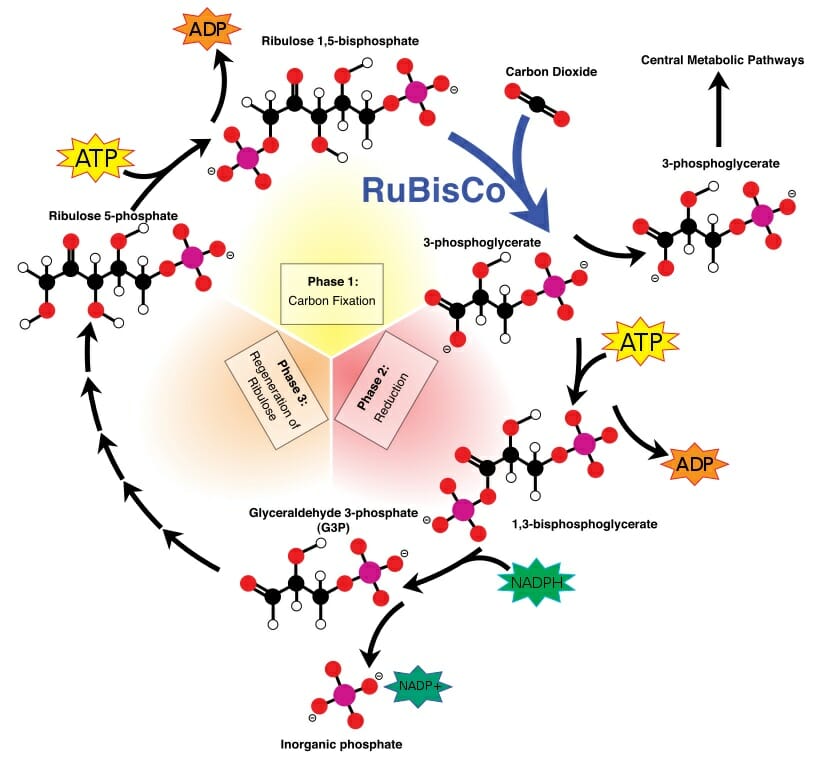
Calvin cycle
Process of Cellular Respiration
Once the glucose is created by the chloroplasts, it can be used to drive other reactions within the cell. It can also be exported to other cells within the organism. This is where the process of cellular respiration takes over. Cellular respiration has 4 distinct processes, which drive the creation of ATP. This ATP can be used in a number of cellular reactions, and provides activation energy to help enzymes complete tasks.
Cellular respiration happens in the mitochondria, a small organelle similar to the chloroplasts. While chloroplasts are only found in plants, mitochondria are found in all living eukaryotes. Plants provide all the glucose their cells need, and more. This extra glucose they store as starches and complex sugars. Animals, and indeed the entire food-chain, relies on the glucose produced by plants.
Cellular Respiration Reaction
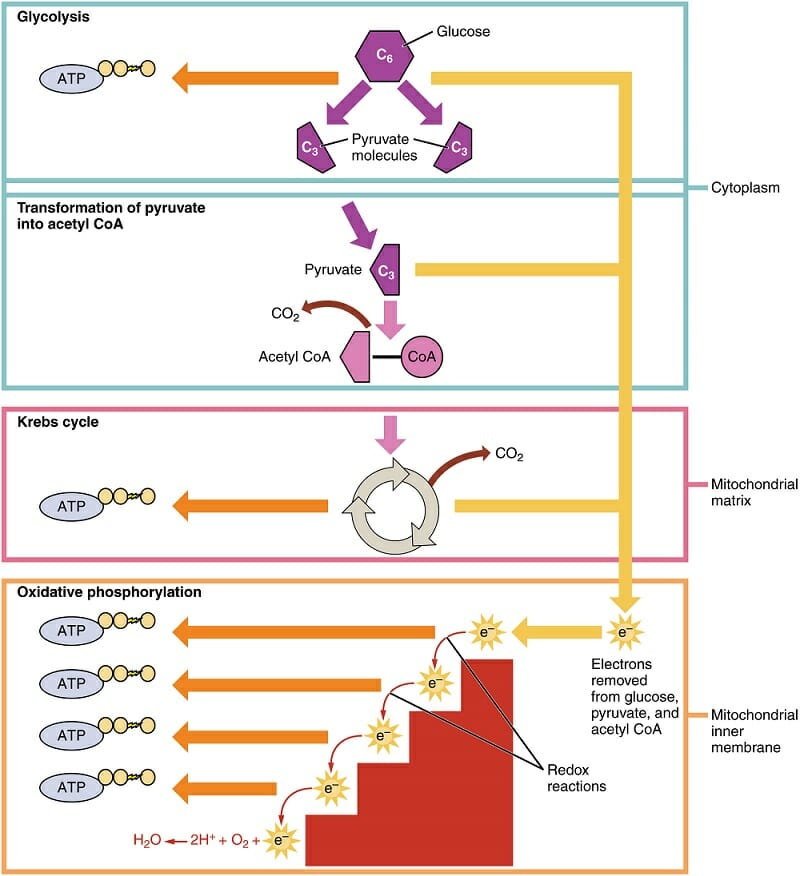
The first process of cellular respiration, glycolysis, is exactly what its name implies. “Glyco-” refers to glucose, where “-lysis” refers to something being divided or split in half. Glycolysis happens within the cytosol of the cell, outside of the mitochondria. In this process, the 6-carbon glucose molecule is split into two molecules of pyruvate.
This 3-carbon molecules is then converted to Acetyl CoA in the next step. This molecule will be an essential part of the Krebs cycle. Acetyl CoA is also able to transfer into the mitochondria, where the Krebs cycle and oxidative phosphorylation will take place. This can be seen in the diagram below. The labels on the right show where the various reactions take place.
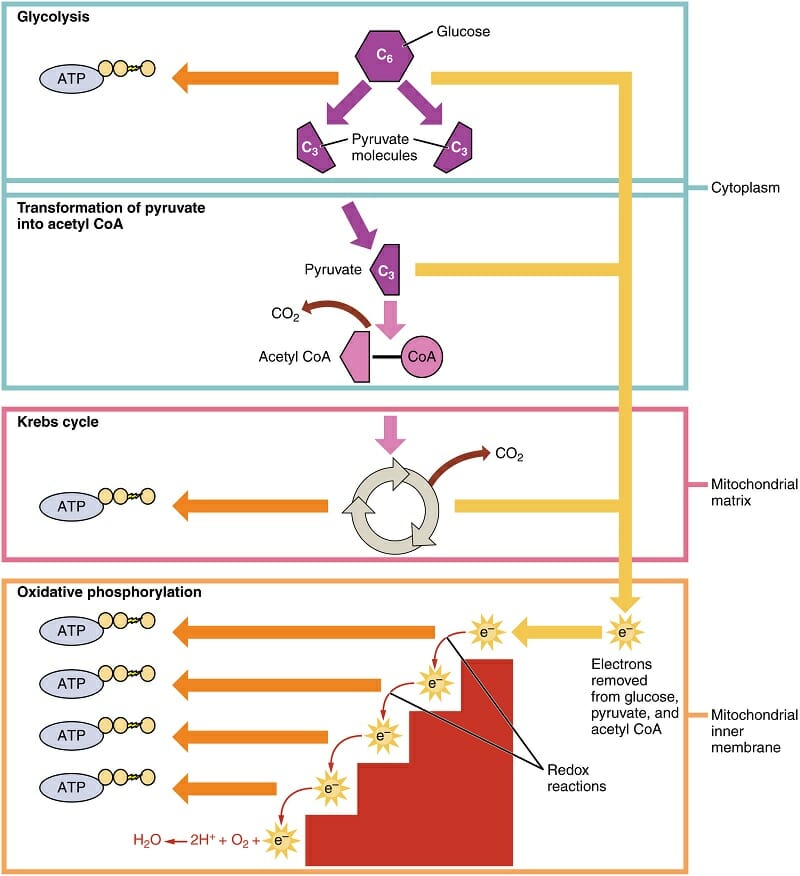
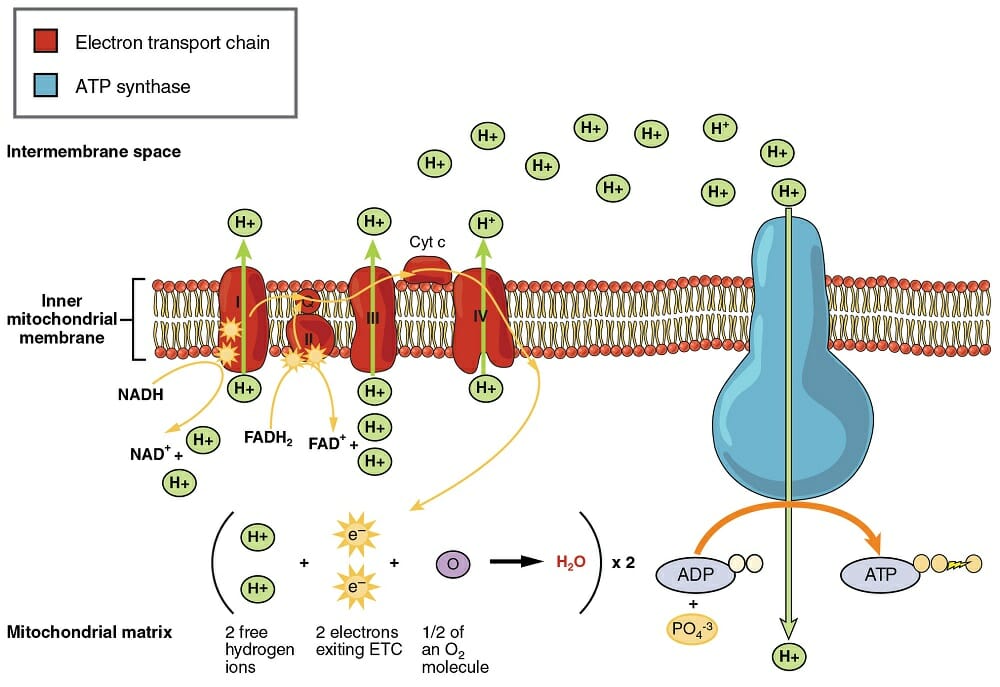
Cellular Respiration
The Krebs cycle is similar to the Calvin cycle, in that it recycles certain molecules to continually drive the production of electrons and ATP. The electrons are then passed to the inner mitochondrial membrane. This membrane is loaded with specialized proteins, capable of transferring energy derived from the passing of electrons down their potential gradient.
This electron transport chain uses a series of electron driven enzymes, which specialize in binding loose phosphate groups to ADP. In doing so, they store energy in the bond between these molecules, and create an ATP. These ATP molecules are then exported from the mitochondria, and can be used throughout the cell to provide energy in other reactions. For instance, ATP is used to pump ions out of cells, creating the electrical potential needed for nervous reactions. There are innumerous other examples.
Cellular Respiration, Photosynthesis, and Evolution
In the Theory of Evolution, the origins of life on Earth are highly unproven. However, there is a large body of evidence which points to the fact that all life has a common ancestor. This ancestor then diverged, over hundreds of millions of years, into the millions of species we see on Earth today. The process of endosymbiosis would account for this complexity.
Bacteria, the simplest organisms, likely represent a fairly unchanged version of the first form of life. Bacteria have no organelles, and complete all the reactions they need for metabolism within a single compartment. Many bacteria are able to complete glycolysis, which can provide them with energy. Others are able to photosynthesize, like primitive single-celled plants.
According to Endosymbiotic theory, these ancient bacteria began interacting and the processes of evolution drove them into different niches within the environment. Some would harness sunlight, while others would feed upon those. Eventually, some of the predatory bacteria became quite large. As such, they could take in large quantities of smaller bacteria. Instead of digesting them, they created a safe space for them and helped them produce more energy. Thus, the smaller endosymbiotic bacteria became the first organelles.
This theory suggests that chloroplasts were originally photosynthetic bacteria, and that mitochondria were originally bacteria capable of oxidative phosphorylation. The larger bacteria became eukaryotes, and developed other organelles. This theory is backed by the evidence that both chloroplasts and mitochondria are surrounded in double membranes, a supposed remnant of the ancestral engulfing process. Further, both mitochondria and chloroplast contain bits of circular DNA, similar to that found in bacteria. This DNA is replicated separately from the main DNA found within the nucleus.
Cellular Respiration, Photosynthesis, and Ecology
Hundreds of millions of years after this division of organelles, and evolution has given us what we see today. Plants are related to algae, which are related to photosynthetic bacteria. Animals are related to the ancient organisms which did not receive photosynthetic endosymbionts, and instead relied on consuming other organisms.
At the bottom of the food-chain sit the photosynthetic organisms. They form by far the largest biomass on Earth, limited only by the amount of sunlight, nutrients, and water they receive. One step above plants and algae, herbivores exploit the bounty that plants produce. Some of the largest animals in the world, such as the elephant, are entirely herbivorous. But, there are herbivores of every size, all the way down to grasshoppers and tiny insects. Because an herbivore must consume many photosynthetic organisms to grow, there are many less organisms on this level of the food-chain.
Likewise, there are many less carnivores than there are herbivores, because they must feed on many smaller organisms throughout their life to grow and reproduce. In this way, the entire food-chain and ecology in general is entirely based off of the processes of photosynthesis and cellular respiration. Ecology is also the study of how various organisms interact with each other while carrying out these reactions.
Quiz
1. Which of the following is NOT a difference between photosynthesis and cellular respiration
A. Only one uses sunlight
B. Only one breaks glucose down
C. Only one relies on a cycle of carbon molecules
Answer to Question #1
C is correct. The Krebs cycle and the Calvin cycle, while differing in their outputs, both rely on a chain of carbon molecules which are continually recycled. The molecules are different, but the processes are very similar.
2. As a human, your cells rely on glucose to function. Where does this glucose come from?
A. Your body
B. Plants
C. Meat
Answer to Question #2
B is correct. All of the food that you consume was at one point a plant. If you eat meat, the nutrients you receive from that meat are the same nutrients that animal ate before it died. Even the protein and fat in animals is simply a reuse of the protein and glucose found in plants.
3. Which of the following things would be MOST devastating to an ecosystem?
A. All the grass in a meadow is killed with an herbicide.
B. All the butterflies in a meadow are killed with a pesticide.
C. All the birds in a meadow are killed by hunters.
Answer to Question #3
A is correct. Without the grass, the entire food chain will collapse. The other two examples represent higher levels of the food chain. Without grass, all the insects would die, and all the birds. But also remember that none of the options are good. Without the birds, insects may eat all the grass and the same result will occur.
References
- Lodish, H., Berk, A., Kaiser, C. A., Krieger, M., Scott, M. P., Bretscher, A., . . . Matsudaira, P. (2008). Molecular Cell Biology (6th ed.). New York: W.H. Freeman and Company.
- McMahon, M. J., Kofranek, A. M., & Rubatzky, V. E. (2011). Plant Science: Growth, Development, and Utilization of Cultivated Plants (5th ed.). Boston: Prentince Hall.
- Nelson, D. L., & Cox, M. M. (2008). Principles of Biochemistry. New York: W.H. Freeman and Company.
Cellular Respiration and Photosynthesis